
Two-Locus Models
The human genome is thought to have about 40,000 genes, Drosophila about 13,000 genes, and even bacteria contain thousands of genes per cell!
Clearly, one-locus models of selection are highly simplified depictions of how populations evolve over time.
Exact analyses with multiple loci are, however, extremely difficult if not impossible to obtain.
Even with only two loci, the dynamics are complicated and not completely understood. Results are limited in scope, focusing on particular fitness schemes.
Nevertheless, results from two-locus models are very important in determining what properties of the one-locus model are unique and might not apply to the real-world situation in which a number of loci collectively interact to guide the formation of the individual.
Consider two loci, A and B, with two alleles each: A1, A2 and B1, B2, at frequencies pA1, pA2 and pB1, pB2.
There are four possible combinations of these alleles on a chromosome:
Chromosome type: A1 B1 A1 B2 A2 B1 A2 B2
Frequency: x1 x2 x3 x4
[Note: x1 + x2 + x3 + x4 = 1]
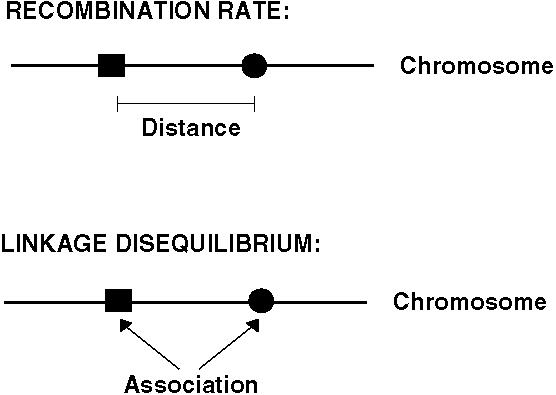
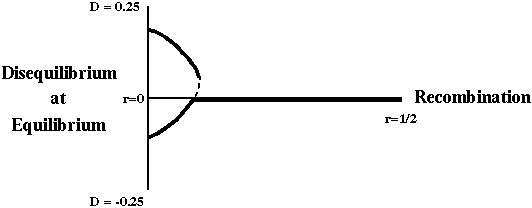
There are two important new concepts in the two-locus model: Recombination and linkage disequilibrium.
Recombination occurs during meiosis in sexual organisms to generate gametes carrying new combinations of alleles. For example:
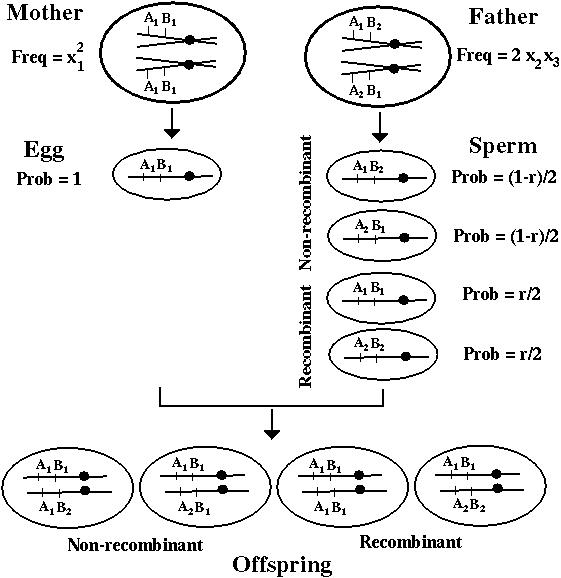
We specify the rate of recombination between two loci by r.
[Note: Recombination may occur in any individual but it only changes the type of offspring produced if the parent was a double heterozygote.]
Linkage disequilibrium, on the other hand, measures whether an allele at one locus is associated (or correlated) with an allele at a second locus.
Linkage disequilibrium, D, is measured by x1 x4 - x2 x3.
Positive D implies that the chromosomes A1 B1 (x1) and A2 B2 (x4) are more common than expected.
Negative D implies that the chromosomes A1 B2 (x2) and A2 B1 (x3) are more common than expected.
Linkage disequilibrium also measures the difference between observed and expected chromosome frequencies:
D = x1 - pA1 pB1
- D = x2 - pA1 pB2
- D = x3 - pA2 pB1
D = x4 - pA2 pB2
Recombination rate (r) is a measure of the distance between two loci and equals the probability that a gamete contains a chromosomal combination not found in the parents.
Linkage disequilibrium (D) is a measure of whether an allele at one locus tends to be found more often with an allele at another locus.
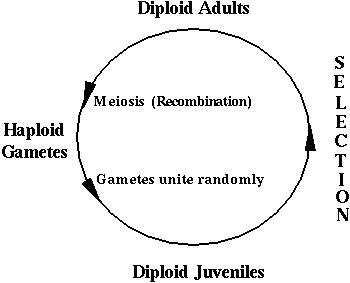
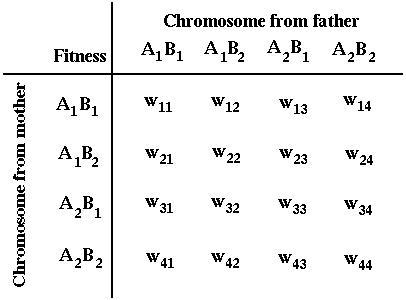
where the survival of a diploid individual depends on its genotype:
These gametes unite at random, creating all possible combinations according to their frequencies. Selection then acts on these diploid genotypes producing a new generation of adults. These adults undergo meiosis to produce the next generation of gametes.
These processes are illustrated in the following mating table:

For instance,
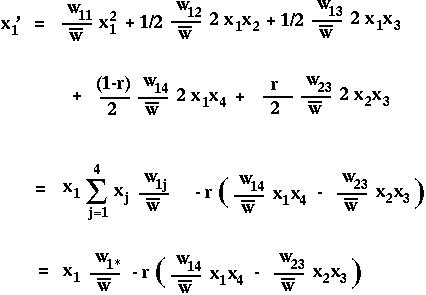
where we define
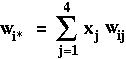
as the marginal fitness of allele i (the fitness that the allele experiences averaged over all genetics backgrounds) and where
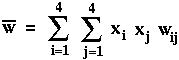
is the mean fitness of all members of the current population.
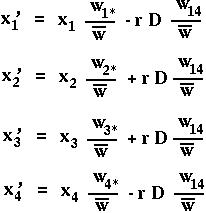
Notice that recombination only enters into these equations when multiplied by the linkage disequilibrium D. (Why is recombination irrelevant if D=0?)
We will not analyse this model in detail, but will simply illustrate some interesting results and how they are obtained.
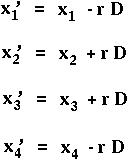
What is p'A1 = x'1+x'2 ?
 In the absence of selection, allele frequencies remain constant.
In the absence of selection, allele frequencies remain constant.
What is D' = x'1x'4 - x'2x'3 ?
Linkage disequilibrium decays at a rate r every generation.
After an amount of time t, the expected amount of disequilibrium is D[t]=(1-r)t D[0].
Therefore, after enough time has passed, we expect to see little linkage disequilibrium between two neutral (=not selected) loci unless they are very tightly linked.
Linkage disequilibrium was measured between several pairs of loci in Drosophila melanogaster.
The statistical evidence for a non-zero D value is here plotted as a function of distance between the pair of loci:
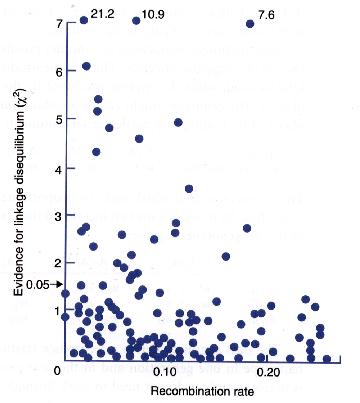
Alleles at most pairs of loci are not significantly associated.
| Assume that the A locus is unselected (neutral), while the B locus experiences selection with fitnesses for B1 B1, B1 B2, and B2 B2 equal to 1, 1+ h s, and 1+s, respectively. | 
|
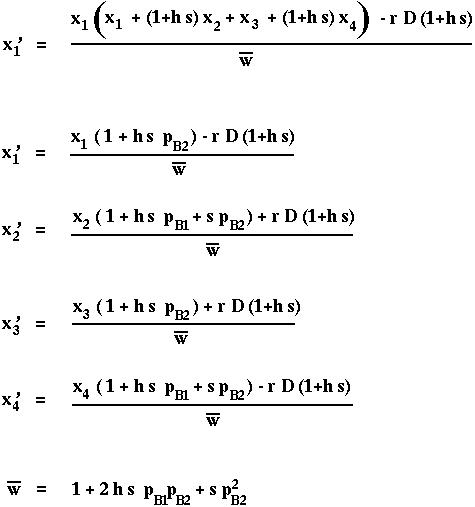
How does the selectively favored allele, B2, change over time?
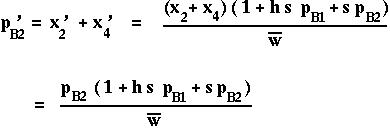
This is identical to the recursion for B2 from a simple one-locus model.
How does the neutral allele, A2, change over time?
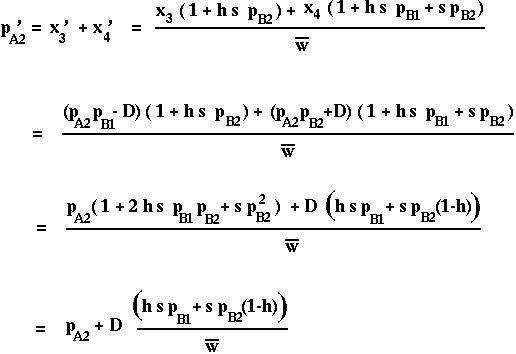
Only when D=0 will the frequency of A2 remain constant over time, as expected from the one-locus model.
Otherwise, the A2 allele will change in frequency because of its association (linkage disequilibrium) with another selected locus: Genetic Hitchhiking.
When s is positive (B2 favored) and D is positive (A1B1 and A2B2 are more common than expected), the A2 allele will increase in frequency over time.
When s is positive (B2 favored) and D is negative (A1B2 and A2B1 are more common than expected), the A2 allele will decrease in frequency over time.
If recombination is loose (r > s), disequilibrium decays rapidly and the A2 allele will change little in frequency.
If recombination is tight (r < s), the extent of hitchhiking can be great.
Implications?
There are, however, some general and counter-intuitive results.
Result 1
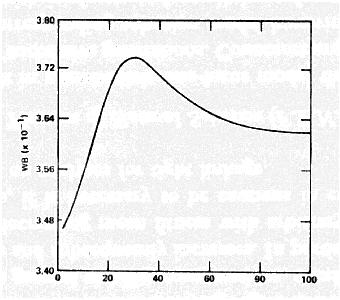
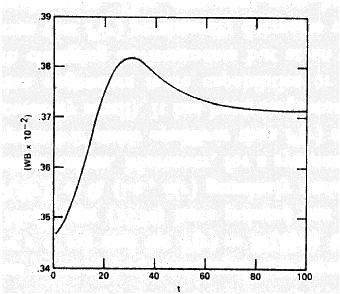
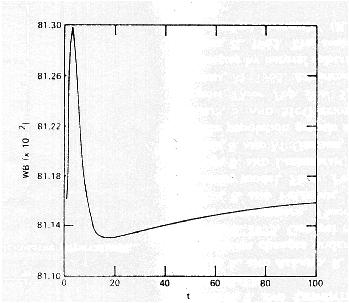
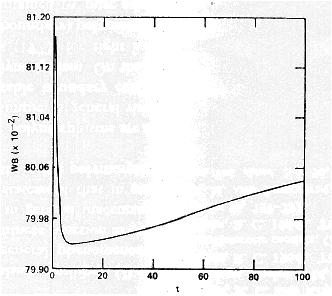
Mean fitness can go down.
For example, Karlin and Carmelli (1975) found that fitness went down for some period of time in most of their simulations of the two-locus model with fitnesses drawn at random:
Result 2
Linkage disequilibrium does not always decay to zero but can be maintained indefinitely within a population.
In fact, very special conditions are required for linkage equilibrium (D=0) to be maintained when both loci are polymorphic and under selection.
Result 3
Continuous cycling can occur (Hastings 1981): "apparent cases of directional selection may be due to stable cycling".
Result 4
Even when the double heterozygote has the highest fitness, there may be no stable equilibrium maintaining polymorphisms at both loci!
Claim 1
If there is pure directional selection favoring a genotype, say A2B2 (such that wi,j+1 > wi,j and wi+1,j > wi,j), it has been claimed but not proven that selection will lead eventually to the fixation of A2B2.
Nordborg (pers. comm.) ran simulations with millions of fitness sets, which all confirmed this claim, but it has not been shown analytically.
More results are known for special cases of the two-locus model:
Special Case 1: r=0
When r=0, the system is dynamically the same as a one-locus four allele model.
Mean fitness never decreases.
Special Case 2: Additive fitnesses
If the fitnesses of the A and B loci add together to give two-locus fitnesses, then:
Special Case 3: Multiplicative fitnesses
If the fitnesses of the A and B loci multiply together to give two-locus fitnesses, then:
IMPORTANT CONCEPTS TO REMEMBER:
SOURCES:
 Back to Zoology 500 home page.
Back to Zoology 500 home page.