
Evolution and Natural Selection

The process of evolution by natural selection rests upon three premises:
(2) Not all individuals survive and certain traits improve fitness.
(3) Traits may be inherited.
 Parents with characteristics that
improve fitness are more likely to have offspring.
If heritable, these characteristics increase in frequency leading the population to change over time (=evolve).
Parents with characteristics that
improve fitness are more likely to have offspring.
If heritable, these characteristics increase in frequency leading the population to change over time (=evolve).
We turn now to the second premise, that selection occurs.
Darwin (1859) in the Origin of Species argued:
"As many more individuals of each species are born than can possibly survive, and as, consequently, there is a frequently recurring struggle for existence, it follows that any being, if it vary slightly in any manner profitable to itself, under the complex and sometimes varying conditions of life, will have a better chance of surviving, and thus be naturally selected."
What factors might contribute to fitness?

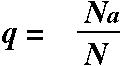
The frequency of A individuals among the offspring is therefore:

Point 1: It is often easier to measure relative fitness rather than absolute fitness.
Point 2: Population genetics models generally require only relative fitnesses, eg
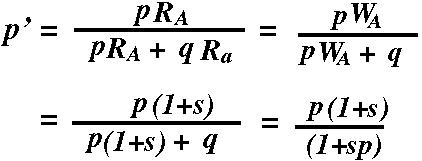
These may be ordered in a number of ways:
More terms to remember:
If WAA=WAa, allele A is said to be "dominant" and allele a is said to be "recessive".
[Note: geneticists usually name alleles that are recessive with lower case letters (eg ubx) and those that are dominant with upper case letters (eg Ubx).]
If WAa=Waa, allele a is said to be "dominant" and allele A is said to be "recessive".
Even more terms to remember:
"Additive" "Partially dominant" "Partially recessive"
Relative fitnesses of AA, Aa, and aa all equal one. Let
x+y+z=1. Why?
Case 1: Individuals produce haploid gametes that form a gamete pool.
The frequency of allele A in the gamete pool will be? p =
The frequency of allele a in the gamete pool will be? q =
Gametes unite at random in the gamete pool to produce diploid offspring. To calculate offspring frequencies we use mating tables.

Point 1: Populations not at Hardy-Weinberg reach Hardy-Weinberg equilibrium after only one generation of random mating (as in the above example). Caveat: Generations must be discrete.
The frequency of allele A in the next gamete pool will be? p' =
The frequency of allele a in the next gamete pool will be? q' =
Point 2: In the absence of selection and mutation, allele frequencies stay constant. Segregation does not change allele frequencies.
Will Hardy-Weinberg frequencies still obtain?
Again, to calculate offspring frequencies we use mating tables.

This shows that x'=p2: Hardy-Weinberg equilibrium is reached after only one generation of random mating.
Since the allele frequency of A in the parents is x+y/2=p (by definition), and since the allele frequency of A in the offspring equals x'+y'/2 = p2 + 2 p q/2 = p (p+q) = p , the allele frequencies again remain constant.
 Back to biology 500D home page.
Back to biology 500D home page.