
Other web sites of interest are:

Sexual selection depends on the success of certain individuals over others of the same sex, in relation to the propagation of the species; while natural selection depends on the success of both sexes, at all ages, in relation to the general conditions of life. The sexual struggle is of two kinds: in the one it is between the individuals of the same sex, generally the males, in order to drive away or kill their rivals, the females remaining passive; while in the other, the struggle is likewise between the individuals of the same sex, in order to excite or charm those of the opposite sex, generally the females, which no longer remain passive, but select the more agreeable partners.The belief in the power of sexual selection rests chiefly on the following considerations: Certain characters are confined to one sex; and this alone renders it probable that in most cases they are connected with the act of reproduction. In innumerable instances these characters are fully developed only at maturity, and often during only a part of the year, which is always the breeding season....Lastly, we have distinct evidence, with some quadrupeds and birds, that the individuals of one sex are capable of feeling a strong antipathy or preference for certain individuals of the other sex.
The Descent of Man and Selection in Relation to Sex (1874 ed., p. 639)
Male as well as female choice has been documented.
We will focus on the more common and better studied cases in which males are more showy and females are more choosy.
Theories for the existence of sexually selected traits
| Horns, antlers, tusks, spurs and other weapons provide some of the most extreme examples of sexual dimorphism. | 
|
Possible explanations:


Eberhard (1979,1980) studied the use of horns "in beetles, finding that they tend to be used either to pry a rival off his site or to lift and drop the rival to the ground.
Brown and Bartalon (1985) placed forked fungus beetles together in a terrarium. Males with the longest horns had higher access to females. Female preference was not observed.
Among the explanations for sexually dimorphic horns, antlers, tusks, and spurs, the empirical support is strongest for the idea that they have evolved and are favored in males as weapons in contests over females.
PHASE 1: Female preferences initially evolve because the preferred trait is favored by natural selection and hence the offspring are more likely to carry the beneficial trait.
Whenever appreciable differences exist in a species, which are in fact correlated with selective advantage, there will be a tendency to select also those individuals of the opposite sex which most clearly discriminate the difference to be observed, and which most decidedly prefer the more advantageous type.
PHASE 2: Once female preferences exist, males with the trait are even more fit (both a natural and a sexual selection advantage). There will then be an ever increasing selective force favoring stronger preferences and more extreme traits (Fisher's runaway process).
...the further development of the plumage character will still proceed, by reason of the advantage gained in sexual selection, even after it has passed the point in development at which its advantage in Natural Selection has ceased.
The runaway process will eventually halt when there is no longer genetic variation for further trait or preference exaggeration or when the viability costs of the trait balance sexual selection.
"Fixed relative preference model", Kirkpatrick (1982):
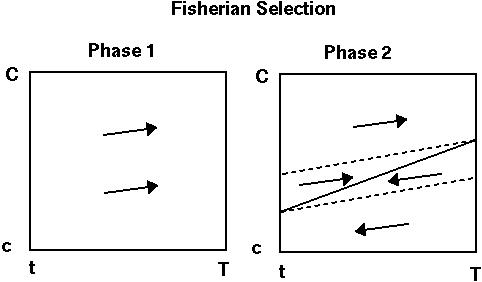
Example:
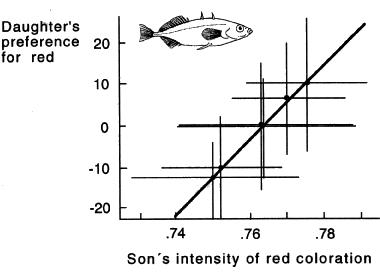
In a breeding experiment with the threespine stickleback, Bakker (1993) observed a genetic correlation between red coloration among sons and preferences for red coloration among daughters, as expected under the Fisherian process.
One main drawback to the Fisherian model of sexual selection is that it is ineffectual in the face of costs to female choosiness:
The converse argument is that when female choosiness is directly beneficial, female preferences will evolve (and the dynamics of the male trait follow).
Possible benefits to being choosy
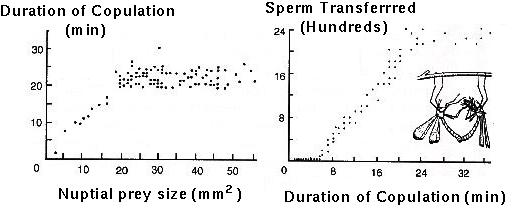
Thornhill (1983) showed that female hangingflies have a higher egg laying rate when mated with males that provide large nuptial food gifts.
| Pleszczynska (1978) showed that male lark buntings with more nest cover in their territory attracted more females, and these females had higher breeding success. | 
|
Linden (1991) showed that great tits avoid previous mates with whom they had few offspring compared with those with whom they had many offspring.
Female preferences can evolve (1) as a correlated response to the evolution of a favored (and preferred) male trait (2) due to direct benefits of choosiness.
An alternative possibility is that female preferences are simply a side effect (a "pleiotropic" effect) of sensory evolution which has evolved for other reasons.
Baker and Carde (1979) showed that males attract females using pheromones normally present in their food.
| Searcy (1992) found that female common grackles preferred males singing an artificial repertoire with four song types even though males in this species sing only one song type. | 
|
Basolo (1990) showed that female platyfish preferred males with swords artificially attached, arguing that evolution in the sister taxa (swordtails) was shaped by this pre-existing bias for swords. [There is debate, however, about the phylogeny; both taxa may have had a common ancestor with swords in which case the preferences do not predate the trait.]
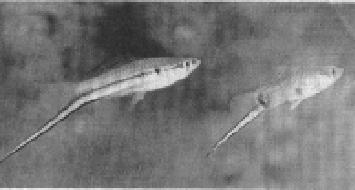
(On the left is a tropical green swordtail, on the right a southern male platyfish with a sword artificially attached.)
 Back to biology 500D home page.
Back to biology 500D home page.