
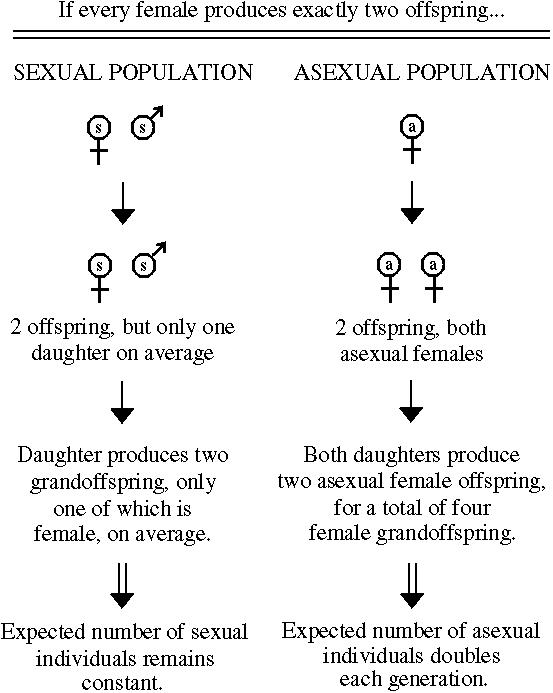
The unit of reproduction is the individual in the case of asexually reproducing organisms but the couple in sexually reproducing organisms.
Unless the sexual couple produces twice as many offspring as the asexual individual, there will be a cost of sex (fewer offspring per sexual parent than per asexual parent).
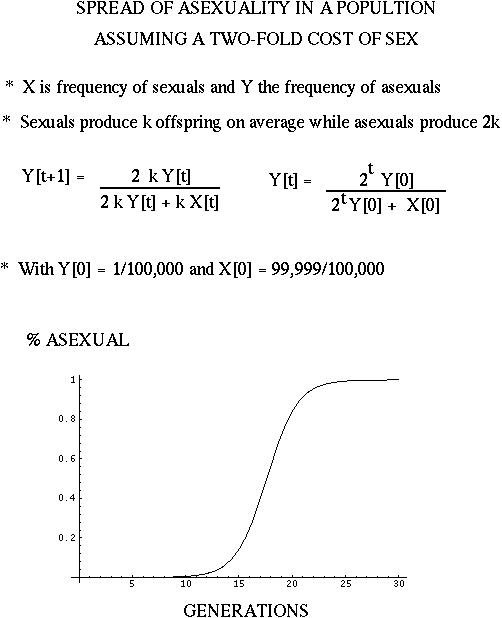
If each female produces a constant number of offspring whether she is sexual or not, then there is a two-fold cost of sex because asexual females produce twice as many female offspring.
In other words, the "sexual female propagates her genome, or any given element of her genome, only half as efficiently as the asexual female" (Bell 1982).
 The Paradox of sexuality
The Paradox of sexuality
Other costs of sex:
What might offset the cost of sex?
What might pay the cost of sex?
In some fashion or other, all explanations of sex rely on the fact that sex can generate greater variability through recombination and segregation.
Sex allows advantageous alleles in different individuals to be combined together into the same individual.
Related:
Also known as the The Vicar of Bray Hypothesis after:
an English cleric noted for an ability to change his religion whenever a new monarch ascended the throne [to emphasize] that there may be great advantages of easily and gracefully adapting to changed circumstances. Bell 1986.
Probability of fixation of beneficial mutations is higher in sexuals with frequent recombination than asexuals with no recombination. The effect substantially favors sexual individuals ONLY if beneficial mutations occur frequently throughout the genome.
| Since the world is constantly changing, organisms must, like the Red Queen in Alice in Wonderland, constantly adapt just to stay in place. |
 |
If sexual organisms adapt faster, they may be able to keep up with rapid changes in the environment, while asexual populations fall behind.
Asexuals may be more prone to extinction.
Parasites are an especially important part of the environment. Organisms must evolve defenses etc continuously and rapidly against viruses, bacteria, insect parasites, etc.
(When focused on the evolutionary arms race" between hosts and parasites, the hypothesis is known as the Host-Parasite Hypothesis.)
Problems with the Fisher-Muller and the Red Queen Hypotheses:
Sex and recombination can break apart new beneficial (e.g. resistant) genotypes once they arise, whereas asexuals can reproduce their genomes exactly.
Mutation and dispersal can sometimes generate and retain more beneficial genotypic combinations than sex.
| If individuals with multiple mutations are less fit than expected based on the effects of each mutation considered separately (i.e. negative epistasis is present), extreme genotypes will become underrepresented in the population (i.e. negative disequilibria will be generated). |
 |
Loss of genetic variation
Sex and recombination can regenerate the extreme genotypes that are underrepresented and increase the amount of genetic variance.
With this increased variability, selection is more efficient at increasing the frequency of the most fit alleles.
(Also known as the Synergistic Epistasis Hypothesis and the Mutation-Load Reduction Hypothesis when considering only deleterious mutations.)
Kondrashov (1988) has argued that if the genomic rate of deleterious mutation (U) is greater than 1, the advantages of sex can outweigh a two-fold cost of sex.
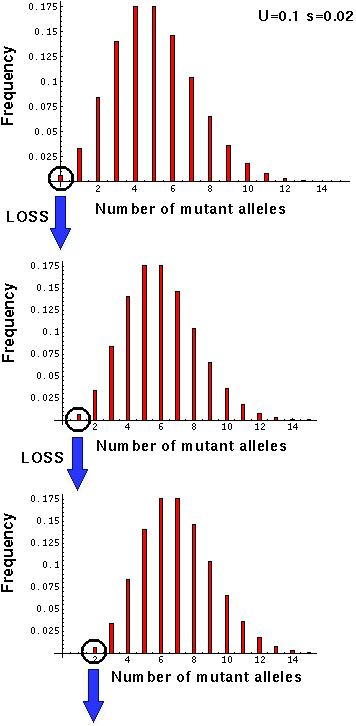
In any population, the fittest genotype may be lost as a result of sampling fluctuations.
Sexual populations can regenerate these lost genotypes by recombination (as long as the alleles have not been lost).
Asexual populations cannot.
In an asexual population, the fittest genotype follows the recursion: p' = (1-
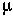 ) p W0/
) p W0/
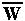 .
At equilibrium, p'=p, so = (1-) W0.
.
At equilibrium, p'=p, so = (1-) W0.
Once the 0th class is lost, the population reequilibrates such that = (1-) W1.
Mean fitness declines, and the average number of mutations increases.
This reequilibration reduces the frequency of individuals in the next best class, which can then be lost, repeating the cycle [= the ratchet].
If we assume that fitness is multiplicative across loci, the number of individuals in the best class (n) equals:
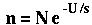
where N is the population size, U is the genomic mutation rate, and s is the selective disadvantage of the mutants.
If we also assume that the number of offspring per parent follows a Poisson distribution, the probability of losing each member of the fittest class is
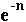 .
.
The ratchet will be fastest when mutation rates are high, selection is weak, and population sizes are small: Greatest advantage to sex under these circumstances.
For instance, with N=100,000, U=0.1, and s=0.01, n=4.5. There is a 1-2% probability that all of 4 to 5 parents fail to produce offspring in any given generation!
The cost of sex could also be lessened if a sexual population were predominantly female.
Fisher (1930) developed a compelling argument for why sex ratios should evolve to 1/2:
The total reproductive value of the males [in a population] is exactly equal to the total value of all the females, because each sex must supply half the ancestry of all future generations of the species...The sex ratio will so adjust itself, under the influence of Natural Selection, that the total parental expenditure incurred in respect of children of each sex, shall be equal. Fisher 1930.If this were not so, parents that produced the rarer sex would gain a larger proportion of that sex's reproductive value and hence a greater genetic representation among grand-offspring.
 Back to biology 500D home page.
Back to biology 500D home page.