PROTOCHORDATES, VERTEBRATE PHYLOGENY AND EMBRYOLOGY

PROTOCHORDATES
Protochordates are an informal category of animals (i.e.: not a proper taxonomic group), named mainly for convenience to describe invertebrate animals that are closely related to vertebrates. This group is composed of the Phylum Hemichordata and the Subphyla Urochordata and Cephalochordata.
The Phylum Hemichordata consists of marine worms that share some, but not all of the characteristics of chordates. These animals have pharyngeal gill slits and a dorsal nerve cord, which is usually solid. The three body parts are proboscis, collar and trunk. What was once thought to be a notochord is no longer considered homologous. Acorn worms are examples of hemichordates.
The Urochordates and Cephalochordates are protochordates, but belong to the Phylum Chordata. Therefore, these animals will be discussed in the chordate section.
CHORDATES
Many characteristics of chordates can only be seen in the embryonic stages. What are the characteristics of this Phylum?
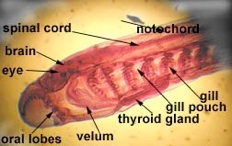
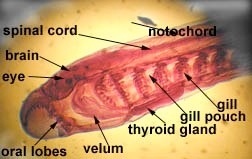
Chordates are subdivided into three subphyla. The subphylum Urochordata consists of the tunicates and related animals. Look at the adult tunicate and you will see very little in the way of Chordate characteristics. This is because the adults have become sessile filter feeders. The pharyngeal gill slits form a large filtering basket and the endostyle (thyroid precursor) produces mucus to trap small particles. If you look at the tadpole larval stage instead of the adult, you will see many more characteristics of the Chordates.
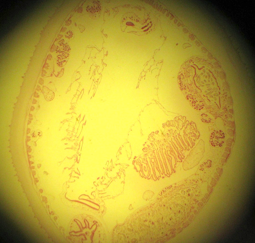
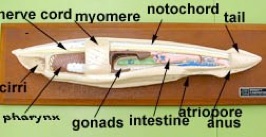
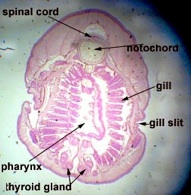
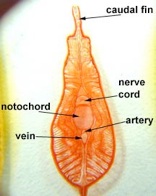
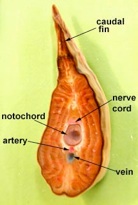
Subphylum Cephalochordata is represented by the little sand lance, Branchiostoma. This marine animal looks very fish-like with fins, a tail, muscle blocks called myomeres and pharyngeal gill slits for filter feeding. Study their anatomy and note their chordate characteristics. The larva looks like a miniature adult without gonads. This is the one group that retains the Chordate characteristics throughout its life.
Branchiostoma
Common name, Amphioxus
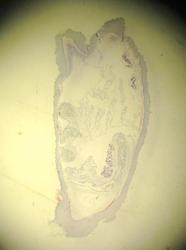
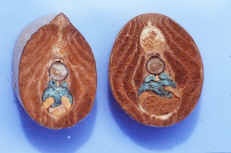
Branchiostoma cross section through the oral hood and through gonads.
Beside this cross-section, draw a Branchiostoma cross-section through the pharynx or gut, showing the gonads.
Subphylum Vertebrata is extremely diverse. The notochord may be replaced partially or entirely by vertebrae, which protect the dorsal nerve cord. They have a three part brain in a brain case with specialized sense organs, a closed circulatory system, a chambered and muscular heart, red blood cells, and glomerular kidneys. This phylum consists of eight classes:
Class Myxinoidea and Class Petromyzontida
Myxinoidea (hagfish) and Petromyzontida (lampreys) are primitive cartilaginous fish, without jaws, called agnatha (a=without, gnatha=jaws) or cyclostomes. We will study the lamprey as a typical vertebrate in the next section.
Hagfish are marine deep water “slime eels.” They have rudimentary eyes, one nostril, one posterior fin, and sucking mouths with two rows of teeth-like projections on the tongue, which they use to eat dead or dying fish from the inside out. Mucous glands produce large quantities of slime, released through pores in the skin, to deter predators. Hagfish have soft, scaleless skin, commercially called eel skin. Inside they have a cartilaginous skeleton. Modern forms lack vertebrae although they are present in fossil forms. They have a muscular body divided into myotomes.
What distinguishes the modern day Myxinoidea (hagfish) from other members of the Vertebrata?
Class Chondrichthyes
This group consists of the sharks and rays (Elasmobranchii) and the chimeras, or ratfish, (Holocephali). As the name Chondrichthyes (‘cartilage’ and ‘fish’) implies, the skeleton of these animals is composed primarily of cartilage. Examine some of the examples on display of the animals in this class. Note the numerous gill slits, the anteriormost of which is sometimes reduced to a spiracle. The tail of sharks and rays is unequal or heterocercal with the upper lobe larger than the lower. Scales are placoid, and oils in the liver keep them afloat. . Internally the intestine has a unique way of increasing digestive area. Instead of folding, it has an internal spiral valve through which the digested food slowly descends.
Class Osteichthyes
These are the bony fish, although a few have reverted to cartilage. Most bony fish have gas filled swim bladders. Most of these fish belong to the Actinopterygii or ray-finned fish. Some are primitive, for example the paddlefish and sturgeon. Somewhat more advanced are the gars and bowfin, Amia. Recent ray-finned fish belong to the Teleosts and this is the largest group of fish. Their skulls have mobile jaws, their gills are protected by a large flap, the operculum, and their tails are homocercal (even sized lobes), and their scales do not have ganoin. The second group of bony fish is the Sarcopterygii and they have fleshy fins with a basal projecting appendage containing bone and muscle. This fin type is thought to give rise to the first terrestrial vertebrates. Here we find the lungfish and the coelacanth. In this group, scales tend to be covered with cosmine and nostrils open internally to the mouth through holes called choanae.
Class Amphibia
The development of a muscular limb with defined joints and digits (tetrapod) is first seen in the amphibians. Also, for the first time an atlas allows the head can move up and down separately from the trunk. Modern amphibia have eggs with no shells or membranes which are laid in water, usually have paired lungs, have mucous glands to keep them moist and poison glands for protection. Amphibia tend to reduce bones, lose scales and are small. They can be divided into three groups, Urodela (salamanders and newts), Anura (frogs and toads) which lack an adult tail and undergo the most dramatic metamorphosis from aquatic tadpoles, and the Gymnophiona (caecilians or apodans) which have no trace of limbs or girdles and are burrowers.
Class Reptilia
Reptiles are the first amniotes, having eggs with extra embryonic membranes, which allow the embryo to develop on land. As we will see in the skull lab, a lot of the relationships are based on the holes in the skull called fenestrae, which are filled with jaw muscles and on the position of the temporal arches. Many reptile groups, for example the dinosaurs, are extinct. The Sauropsids, which includes the dinosaurs, contains the modern day reptiles and led to the birds. Another group, the Synapsids, produced the extinct therapsid reptiles and led to modern mammals.
Modern reptiles with no temporal fossa (anapsid) are the Parareptilia (turtles). Most modern reptiles have two temporal fossae (diapsid) and include snakes, lizards, legless lizards, crocodiles, alligators and the birds. A second neck bone, the axis, allows greater mobility of the head and most reptiles have long necks with many cervical vertebrae.
Class Aves
Birds are specialized diapsid reptiles, which are second only to fish in number of vertebrate species. Their closest relatives are the crocodiles; both lay shelled eggs and have similar musculature and bones, including a wishbone. Birds are warm blooded, have feathers and complex lungs. Birds usually have wings for flying and hollow, air-filled bones which keep them light. They also have beaks and walk on two legs (bipedal).
Class Mammalia
The mammalian skull, like that of the therapsid reptiles has only the lower temporal fossa and thus is a synapsid skull. Mammals are warm-blooded, have hair and mammary glands. They also have sebaceous (oil) glands and sweat glands. They have anucleate red blood cells. Mammals have three bones in the middle ear and most mammals have large brains. The lower jaw is composed of a singe bone, the dentary, which forms a joint with the squamosal bone. Although the dentaries are paired structures, they are often fused into a single structure by the mandibular symphysis.
There are three groups of living mammals. The monotremes include the duckbill platypus and spiny anteater. They are amniotes with a primitive, shelled egg. Another group of mammals, represented by the opossum and best known by the kangaroo, are the Monotheria (marsupials). These animals have a simple placenta and young born at a very early stage, which crawl into a pouch and suckle there until they are larger. Most modern mammals are Eutheria (placentals), with the fetus connected to a complex placenta in the uterus enabling it to develop considerably before it is born.
LAMPREY AS A SIMPLE VERTEBRATE
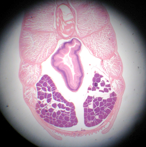
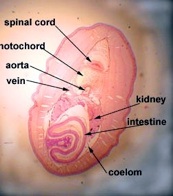
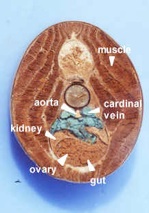
Lampreys are parasitic, clinging to fish and sucking their blood by rasping away skin using the teeth on the tongue. Since blood is easy to digest, they are all gut (no stomach), which exits in a common cloaca with the kidney and gonads. Like hagfish, lamprey have a single nostril, a cartilaginous skeleton (with vertebrae), and a muscular body divided into myotomes. There are whole mounts of lampreys and cross sections on display.
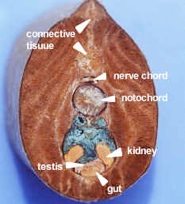
There are a series of lamprey cross sections of the trunk region cut posterior to the heart in plastic mounts and bowls on the bench. View using a dissecting microscope. The liver is anterior to the gonads. These animals are dioecious (separate sexes) and the gonads are small in the males (testes), but large and pebbly in the females (ovaries). The gut is a tiny hollow tube with endoderm inside and mesoderm outside. The kidneys are paired and large, blue on the top because of the injected blood supply. The tail has no digestive or reproductive parts but the muscles, nerve cord and notochord continue.
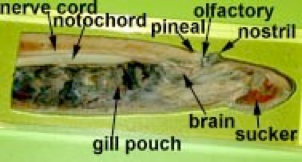
The larval lamprey always lives in fresh water where it burrows and filter feeds. It is called an ammocoetes larva and bears a strong resemblance to the sand lancet. Examine the specimens on slides in the lab. Unlike Amphioxus, the pharynx has muscles and cartilaginous bars so it is capable of pumping action. Other derived characters are the presence of a liver, gall bladder and heart as well as pronephric (kidney) tubules. The brain, even in the larva, is composed of three parts (forebrain, midbrain and hindbrain). After 3-7 years the larva metamorphoses into a juvenile and migrates to the sea or in freshwater species to lakes to take up its parasitic existence.
EMBRYOLOGY
We can classify animals based on the type of development they undergo. Vertebrates have deuterostome development. What does the word deuterostome mean? What are the characteristics of this type of development?
Vertebrates are not the only animals with deuterostome development. We presume that other phyla with this type of development are closely related. The Phylum Echinodermata (starfish, urchins, sea cucumbers, feather stars etc.) is an invertebrate phylum but if we look at their early embryology, it is considered the same. Most other invertebrates undergo protostome development.
Compare deuterostome development with protostome development.
In this laboratory session we are going to study the basic plan of the vertebrate embryo. Four species are compared: models of the sea urchin show the basic pattern of deuterostome development. The embryos of the urchin and Amphioxus, the lancelet, are without, or have very little yoke (microlecithal), having holoblastic cleavage and the coelom is formed by a pouched-off mesoderm (enterocoely). The frog embryo has yolk (mesolecithal), meroblastic cleavage and the coelom forms by a splitting of the mesoderm (schizocoely). The chick embryo has a large-yolked (macrolecithal) egg.
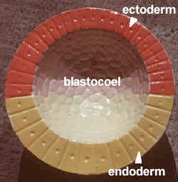
Convention gives three colors to the germ layers. The ectoderm is traditionally colored blue in embryos, the mesoderm is red and the endoderm is yellow. It may help if you color your lab handouts in the traditional colors. Unfortunately, the embryo models that you will study in this lab are not of the conventional colors, so we have given you the correct colors in brackets.
Amphioxus models
Examine the models of Amphioxus development series 1 to 23 on demonstration. Review early embryology starting with the fertilized egg (zygote) and advancing through the various stages in development. The morula (little berry) is an almost solid ball of cells whereas the blastula is a hollow ball of cells.
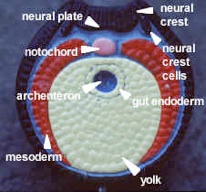
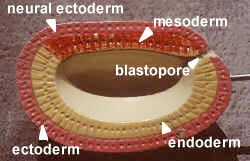
In model #12 the gastrula (gaster = gut or stomach), the top inner layer of mesoderm is called the chordamesoderm because it induces the notochord. As the cells roll into the dorsal lip of the blastopore they become mesoderm. The pink on the model is ectoderm (blue). The yellow endoderm forms the archenteron or primitive gut.
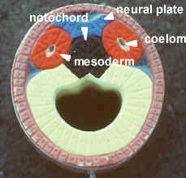
Model #17 shows neural plate formation (see figure on this page). The black mass is the notochord (red). The coelom is pouched off (enterocoely) from the archenteron. Later, organs will hang in the coelomic space. The grey represents mesenchyme, embryonic tissue from mesoderm (red) with unfixed potential to differentiate. It is not a primary germ layer but in the embryo is amoeboid and can become any layer. It is like adult connective tissue which makes up the bulk of the body. Since changes move from anterior to posterior, the oldest part of the section is anterior.
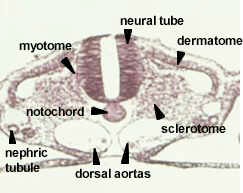
Model #22 shows a dorsal, tubular nerve cord. The mesoderm grows in between the endoderm and ectoderm, eventually forming mesodermal somites. The muscle mass of the somites, epimere myotome, is purple-grey, next to the notochord. The inner layer of hypomere mesoderm (splanchnic) forms the outer wall of the gut and the outer layer of hypomere mesoderm (somatic) lines the body wall. The neural crest cells are absent on this model. They are segmented and run the length of the body to form the branchial skeleton and sensory nerves. The right side of the model shows a slightly more developed view with a pinched-off mesodermal space in the somite.
Frog models
Examine the models of frog development (series 1 to 15), noticing the influence of yolk on cleavage patterns. The neural crest cells are shown in these models. The notochord (red) is painted pink and the ectoderm (blue) is painted brown. Note the gut buried in the yolk. The mesoderm grows as a sheet, the coelomic cavity appearing later by schizocoely, which has been secondarily derived. Segments only appear in the dorsal region.
Chick embryo slides
There are demonstration slides of chick whole mounts and cross-sections on the side bench. A chick has discoidal cleavage in which the yolk does not divide.
In the whole mount, the somites (or myotomes) are repeated blocks of muscles. In this view you see epimere mesoderm.
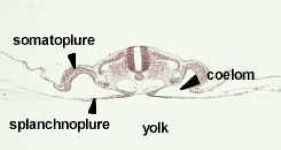
The chick cross section is somewhat like the adult germ layer structures in your handout, but the drawings posted in the lab are better. The chick is sitting on top of the yolk and the gut is open to the yolk. Note the slit-like coelom. The mesoderm is composed of the three parts drawn below (epimere, mesomere and hypomere).
Transverse section through the trunk region of a 33 hour chick embryo showing the establishment of the typical chordate plan and the differentiation of the mesoderm
In the following labs, it will be important to know which adult structures are formed from the three embryonic germ layers (chart pg. 20). Study the chart of the embryonic origins of adult structures. We will be looking at these structures in detail in future labs, but you should always be aware of their germ layer origin. See the chart in your lab manual.