Chapters
Lodish 4th edition: Chapter 21 pages 921 - 924
Moyes and Schulte: Chapter 5 pages 146-164
Active Conduction
- passive conduction of a signal in a nerve is limited by the properties of
the nerve
- not very efficient if need to have signal travel quickly over a long
distance
- signal is reduced over distance
- active conduction (i.e. generating an action potential) means signal travels
along nerve with NO loss of amplitude
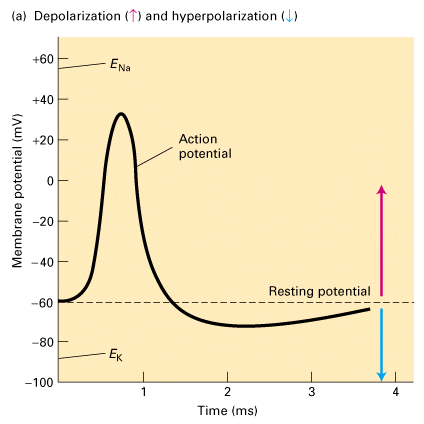

Figure 21-2. Lodish 4th edition (a) An action potential is a sudden, transient depolarization of the membrane followed by repolarization to the resting potential of about -60 mV. This recording of the axonal membrane potential in a presynaptic neuron shows that it is generating one action potential about every 4 milliseconds. (b) The membrane potential across the plasma membrane of a presynaptic neuron is measured by a small electrode inserted into it. Action potentials move down the axon at speeds up to 100 meters per second. Their arrival at a synapse causes release of neurotransmitters that bind to receptors in the postsynaptic cell, generally depolarizing the membrane (making the potential less negative) and tending to induce an action potential in it.
The Channels of an Action Potential
Action potentials in neurons are mostly based on the voltage-gated Na+ channel, some neurons use both the voltage-gated Na+ channel and a voltage-gated K+ channel, some neurons use only the voltage-gated Na+ channel and some neurons use the voltage-gated Ca+2 channel
We will use the classic example of an action potential from the giant axon of the squid (invertebrate), also the action potential found in non-myelinated axons of mammals. This action potential has two components: voltage-gated Na+ channels and voltage-gated K+ channels
Voltage gated Na+ channel:
The channel has three states, closed, open and inactive.
Closed to Open: Depolarization is necessary to open the channel and
therefore it acts to activate itself in a regenerative cycle. More Na+ influx
depolarizes the membrane which opens more channels which depolarizes the
membrane more.
Open to Inactive: Depolarization is also necessary to inactive the
channel. Once the channel is open it will then also switch to the inactive
state and can not be opened again
Inactive to closed: The channel will not switch back to the closed
state until the membrane has repolarized (i.e. gone back towards the original
resting membrane potential. Once in the closed state it can then be reopened
Voltage-gated K+ channel (called the delayed rectifying K+ channel)
This channel has only two states, closed and open.
Closed to open: The channel is opened with a strong depolarization, the
type you would normally get in an action potential. This channel works to
bring the membrane back towards the Nernst potential for K+ i.e. hyperpolarize
the membrane
Open to closed: The channel will close when the membrane becomes
hyperpolarized or repolarized. Therefore this channel works to shut itself
down.
The components of an action potential
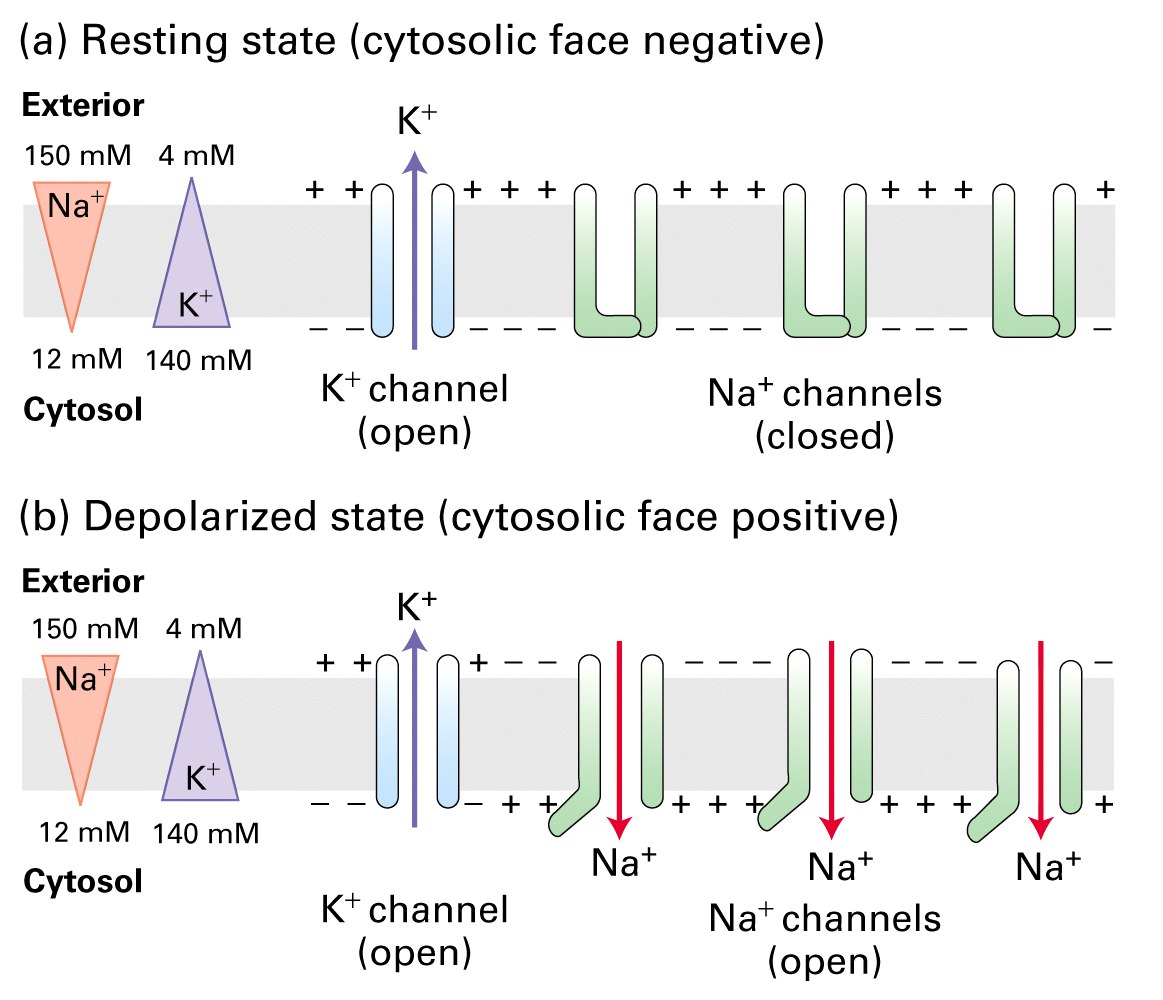
Figure 7-32 Lodish 5th edition. Depolarization of the plasma membrane due to opening of gated Na+ channels. (a) Resting neurons non gated K+ channels are open, but the more numerous gated Na+ channels are closed. The movement of K+ ions outward establishes the inside-negative membrane potential characteristic of most cells. (b) Opening of gated Na+ channels permits an influx of sufficient Na+ ions to cause a reversal of the membrane potential.
Threshold
- level of depolarization needed to trigger an action potential (most
neurons have a threshold at -50 mV (i.e. 10 to 15 mV depolarization)
- an action potential is an all or none event, if a nerve is at rest the
amplitude on one action potential will be the same all along the nerve
independent of the stimulus strength.
- threshold reflects the need to trigger the opening of the voltage-gated
sodium channel (need a depolarization of about 10 to 15 mV to open)
Action potential rising phase
- as sodium channels open, Na+ ions flow into cell, depolarizes the cell more and more sodium channels open = a regenerative response - regenerative opening of sodium channels drives the membrane potential towards a peak of the Nernst equilibrium potential for Na+
Peak of Action Potential
- during an action potential the membrane potential goes towards the Nernst
equilibrium potential for Na+
- in terms of Goldman-Katz equation now permeability to Na+ is dominant (K+
and Cl- minor components) therefore membrane potential goes towards ENa
- usually falls short of ENa, less driving force on Na+ and the channels begin
to inactivate rapidly after activation
Falling Phase of Action Potential
- after reaches peak now action potential falls, membrane potential falls
back towards rest
- why? Why doesn't the action potential stay around ENa?
- two reasons:
i) Na+ channels move into an inactive state
ii) delayed K+ channels open (giant axon of squid or non-myelinated axons of
vertebrates)
1) Inactivating Na+ channels -
- Na+ channels go to an inactivated state after 1-2 msec after first opening
- inactivated = can NOT be reopened
- therefore the membrane potential now determined mostly by K+ (same as for resting potential) and membrane starts to repolarize
2) Delayed K+ channels open (called delayed rectifier; voltage-gated like Na+ channel)
- open after about 1-2 msec of threshold depolarization
- now K+ flows out of the cell and speeds the repolarization process
- cause the hyperpolarization after the action potential because open K+ channels make the K+ permeability higher than at rest and membrane more negative on inside-hyperpolarization of membrane causes K+ channels to close
-then membrane settles back to rest
Repolarization
- voltage-gated Na+ channels and voltage-gated K+ channels now closed so
the membrane goes back to the resting state
- i.e. the leak channels are the only channels open and again set the membrane
potential
Refractory period
-divided into two parts
i) absolute refractory period
ii) relative refractory period
1) Absolute refractory period
- Na+ channels are inactive and CAN NOT be opened no matter how much the
membrane is depolarized at this time
- another action potential can not be generated in this part of the nerve at
this time
2) Relative refractory period
- as membrane repolarize's = goes to more negative potentials this triggers
the Na+ channels to move from an inactive state to a close state.
- hyperpolarization by the opening of the K+ channels helps this process
- once Na+ channel is in the closed state can be opened again with
depolarization
- during relative refractory period, more and more Na+ channels available to
be opened and therefore increase the chances of firing an action potential
Frequency of Action Potentials
- If the action potential if all or none how does a nerve convey the
strength of a stimulus?
- e.g. how does a sensory nerve distinguish between a light touch (feather)
and a rough abrasive touch (sand paper)?
- the information is indicted by the frequency of the action potentials along
the nerve.
-the stimulus strength (current input in to the nerve either experimentally by
injecting a large current or in real life by response of touch receptor)
triggers different frequency of action potentials
-therefore: light touch - infrequent action potentials; rough touch - more
frequent action potentials
- the refractory period limits the frequency of the action potential
- during the relative refractory period an action potential can be generated
but with an increased threshold and a reduced amplitude
- increased threshold because have to over come hyperpolarization
- decrease amplitude because less Na+ channels are available to open (many are
still in the inactive state) and so get less Na+ flowing into the cell
(in other words the permeability or conductance of Na+ is reduced during
relative refractory period - increases towards the end of the period)
Direction of Action Potentials
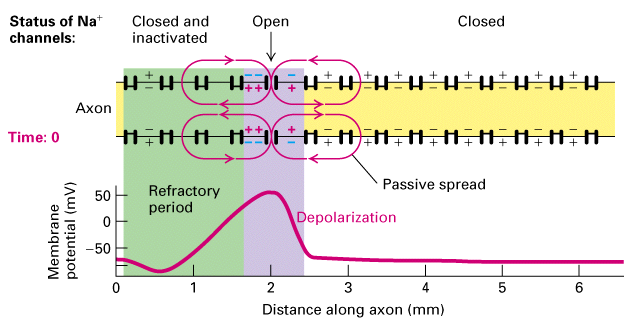

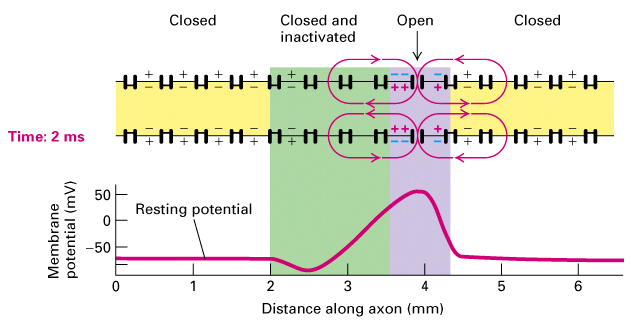
Figure 21-14, Lodish 4th edition OR Figure 7-35, Lodish 5th edition. Unidirectional conduction of an action potential due to transient inactivation of voltage-gated Na+ channels. At time 0, an action potential (purple) is at the 2-mm position on the axon. The membrane depolarization spreads passively in both directions along the axon (Figure 21-11). Because the Na+ channels at the 1-mm position are still inactivated (green), they cannot yet be reopened by the small depolarization caused by passive spread. Each region of the membrane is refractory (inactive) for a few milliseconds after an action potential has passed. Thus, the depolarization at the 2-mm site at time 0 triggers action potentials downstream only; at 1 ms an action potential is passing the 3-mm position, and at 2 ms, an action potential is passing the 4-mm position.
- the refractory period also sets the direction of an action potential
- depolarizing current from the action potential can spread passively in
either direction
- one way the Na+ channels are in a closed state and are ready to be opened,
therefore the spreading current can trigger an action potential in this
neighbouring region
- the other way the Na+ channels are in an inactive state and can not be
opened therefore the spreading current has no effect on the channels in this
region and an action potential is not trigger
Channels and receptors
Throughout the lectures we will be introducing a wide range of ion channels. These range from leak channels (K+, Na+, Cl- etc.), voltage-gated ion channels (K+, Na+ and Ca+2 etc.) and aligned gated ion channels (K+/Na+, Cl- etc.).
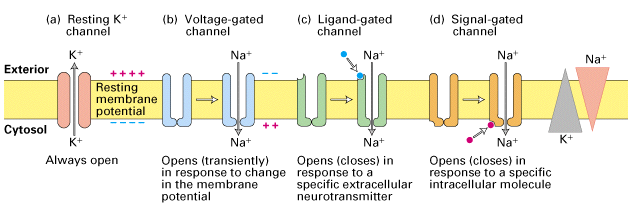
Figure 21-8, Lodish 4th Edition. Ion channels in neuronal plasma membranes.
Each type of channel protein has a specific function in the electrical activity of neurons. (a) Resting K+ channels are responsible for generating the resting potential across the membrane. (b) Voltage- gated channels are responsible for propagating action potentials along the axonal membrane. (c, d) Two types of ion channels in dendrites and cell bodies are responsible for generating electric signals in postsynaptic cells. One type (c) has a site for binding a specific extracellular neurotransmitter (blue circle). The other type (d) is coupled to a neurotransmitter receptor via a G protein; it responds to intracellular signals (red circle) induced by binding of neurotransmitter to a separate receptor protein (not shown). Signals activating different channels include Ca2+, cyclic GMP, and the Ga subunits of trimeric G proteins (Chapter 20).
Most of the proteins that make up the different types of ion channels are very similar in their structure and have conserved amino acid sequences. This degree of conservation occurs between different types of channels and across species. So for instance the Drosophila voltage-gated Na+ channel is very similar to the human voltage-gated Na+ channel etc. All the ion channels are composed of alpha helices that span the lipid bilayer. Those that contact the lipid bilayer are composed of hydrophobic amino acids (Phe, Ile, Leu etc.) that span about 20 amino acids. Those alpha helices that line the pore are composed of hydrophilic residues to allow ion flow (Lys, Arg etc.).

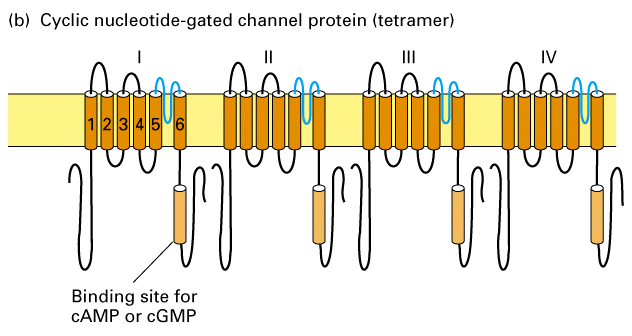

Figure 21-27, Lodish 4th edition. Proposed transmembrane structures of four types of gated ion-channel proteins.
(a) The Shaker voltage-gated K+ channel isolated from Drosophila is a tetramer of four identical subunits each containing 656 amino acids and six transmembrane alpha helices (indicated by Arabic numerals). Helix 4 (maroon) acts as a voltage sensor, and the nonhelical P segments between helices 5 and 6 line the ion pore. (b) Cyclic AMP- or cyclic GMP-gated ion channels also contain four subunits. Since none of the transmembrane alpha helices act as a voltage sensor, these channels are not voltage-gated. Rather, binding of cAMP or cGMP to a cytosolic segment triggers opening of these channels, which are abundant in the sensing cells of the visual and olfactory systems (see Figure 21-47). (c) Voltage-gated Na+ (and Ca+2) channels are monomeric proteins containing 1800 2000 amino acids organized into four homologous transmembrane domains (indicated by Roman numerals). About 64 percent of the residues are similar or identical in sequence in both channels. Each of the four homologous domains is thought to contain six transmembrane alpha helices similar in structure to those in the Shaker channel monomers in (a).
Channel pore
All the ion channels in question have a common feature. A pore that allows the ion(s) in question to flow across the lipid bilayer. The pore is specific to a certain ion or ions. For instance the leak K+ channel only allows K+ ions to flow across the membrane.


Figure 7-16, Lodish 5th edition. Mechanism of ion selectivity and transport in resting K+ channels. (a) Schematic diagram of K+ and Na+ ions hydrated in solution and in the pore of a K+ channel. (b) High-resolution electron-density map obtained from x-ray crystallography showing K+ ions passing through the selectivity filter.
Voltage-sensor
The voltage sensor is an alpha helix is found in the channel and spans the membrane. The voltage-sensor has positive charges at every third amino acid. The sensor moves in response to depolarization (i.e. the increase positive charge on the interior membrane causes the physical movement of the voltage-sensor). The sensor alpha helix is buried within the channel protein (i.e. protected from hydrophobic lipid bilayer by the rest of the ion channel protein).
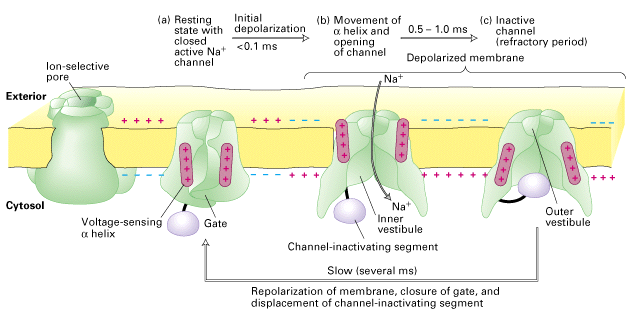
Figure 21-13 Lodish 4th edition OR Figure 7-33 Lodish 5th edition. Structure and function of the voltage-gated Na+ channel.
Like all voltage-gated channels, it contains four transmembrane domains, each of which contributes to the central pore through which ions move. The critical components that control movement of Na+ ions are shown in the cutaway views. (a) In the closed, resting state, the gate obstructs the channel, inhibiting Na+ movement, and the channel-inactivating segment is free in the cytosol. The channel protein contains four voltage-sensing alpha helices (maroon), which have positively charged side chains every third residue. The attraction of these charges for the negative interior of resting cells helps keep the channel closed. (b) When the membrane becomes depolarized (outside negative), the voltage-sensing helices move toward the outer plasma membrane surface, causing an immediate conformational change in the gate segment that opens the channel for influx of Na+ ions. (c) Within a millisecond after opening, the voltage-sensing helices return to the resting position and the channel inactivating segment (purple) moves into the open channel, preventing further ion movements. When the membrane potential is reversed so that the inside is again negative, the gate moves back into the blocking position (not shown). After 1 2 ms the channel-inactivating segment is displaced from the channel opening and the protein reverts to the closed, resting state (a) where it can be opened again by depolarization.
The voltage-sensors move in response to depolarization to open the ion
channel. One model of how the voltage-sensor works is based on a twist or
spiral movement that cause the alpha helix to move within the membrane.
Experimenters can measure this movement of the voltage-sensory but you'll have
to wait until Biology 455 to learn all about that.
The following is a movie of one model of how the voltage-sensor moves.
(WARNING: a big file!!).
Movie of moving voltage-sensor