Biology 303
sec.102, Sept.-Dec. 2004; Some
Thoughts on R0 and
r.
Generally, a
population grows as (Krebs, p.140):
![]()
where: e is a constant
(approx. 2.71828, the base of natural logarithms)
N
is population density
t
is time (0 at the start,
and t some time-period “t” later)
r
is the intrinsic rate of population increase under prevailing conditions
If you wish to
know how quickly a population grows per generation, use G (generation
time):
![]()
(this is given as
“T” instead of “G” in Krebs’ notation)
Take the natural
log of this equation, rearrange it, and you get:
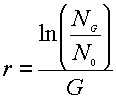
The value in the
brackets is essentially the multiplier by which population size has changed in
one generation, or the relative change in number of breeders per generation. We
also call this term the population net reproductive rate, and symbolize
it as R0.
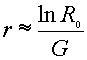
[This value can
also be derived from life-table data; see downloadable Demography document.]
This gives only
the approximate value for r, since it
assumes non-overlapping generations.
The theoretically
ideal value for r – a population’s theoretic intrinsic maximum capacity
for growth – can never be realized by organisms in the real world. An exponentially
growing population in a laboratory container may approach the
theoretical maximum, or rmax, early in its growth, but never be actually “at” it, because as soon as
some individuals are present in an environment that place can no longer support
idealized growth – food has been depleted, waste-products are present, etc.,
it’s no longer a “perfect” growth medium. A population will exhibit a realized situation-specific
value of r, called ra or robs (“actual”, or “observed”, r), which varies
from a little less than rmax, through positive values (increasing population), and zero (stable
population size), to negative values (declining population size). See Box 11.1,
page 162, in Krebs for a different explanation of this point. The units
of r are individual/individual/time, and it’s an instantaneous
rate, saying how the population is changing “right now”.
The instantaneous
value of r is the difference between the per-individual birth
rate and the per-individual death rate at that given instant in time:
![]()
How can rates be
“per individual”? For births, you can imagine the number of offspring
produced per female per season, or in a population in which not all females
breed every season you could have the fraction of total females present
which produced an offspring per season. Either of these is in units of
individual/individual/time.
For deaths, the easiest way to picture it is the number of
individuals dying out of the total number of individuals present during a
time-period, or equivalently the fraction dying per time period.
This sort of measure will also be in units of individual/individual/time.
If you want a finite
rate (a prediction of what will happen a finite period into the future), use
the calculation of l (lambda):
![]()
Krebs offers an
example of this difference in usage at the end of Box 10.2, p.145, but you will
not be asked to use this calculation.