
Biomathematics Practise Questions
Question 1:
(a) C[t+1] = C[t] - 50
(b) No
[Note - During gamete production, a specific enzyme, telomerase, adds to the ends of the chromosomes, without which our chromosomes would eventually disappear.]
Question 2:
(a) p[t+1] = (1-d) p[t] + m/p[t]
(b) Setting p[t+1] = p[t] = 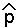 , we get 2 = m/d so there are two equilibria: = Sqrt(m/d) and = - Sqrt(m/d)
, we get 2 = m/d so there are two equilibria: = Sqrt(m/d) and = - Sqrt(m/d)
(c) Only the first equilibrium, with positive, is biologically valid because p represents the level of a gene product, which cannot be negative.
(d) Calling the recursion equation f(p), df/dp = (1-d) - m/p2, which equals 1 - 2 d at = Sqrt(m/d). Under the assumption that d represents the fraction of gene products that decay per time step and must lie between 0 and 1, 1 - 2 d will be less than one in magnitude. Thus, the equilibrium will be stable. If the decay rate is high, d>1/2, then the system will oscillate towards the stable equilibrium. If the decay rate is low, d<1/2, then the system will smoothly approach the stable equilibrium.
Question 3:
(a) f = 1/2
(b)
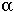 dt
dt
t + c
t + c
t + c]
t + c]
t] (1/2 - f0)
Question 4:
(a) dI/dt = 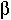 - I
- I
(b) /
(c) Let f = dI/dt, then df/dI = - . For the equilibrium to be stable, df/dI evaluated at the equilibrium must be negative. But df/dI = - for all values of I including at the equilibrium. Therefore, as long as is positive and insulin molecules do degrade over time, this equilibrium will be stable.
(d)
- I) = dt
- I)]/ = t + c
: Ln[( - I)] = - t - c
- I) = Exp[- t - c]
c] = ( - I0)
- Exp[- t] ( - I0))/
Question 5:
(a) We can write a recursion equation using a model of selection with two types, hawks and doves, where p[t+1] = p[t]*Wh/(p[t]*Wh+(1-p[t])*Wd) is a recursion for the frequency of hawks. This is equivalent to the haploid model, but now the two type are hawks and doves, rather than A and a alleles. Plugging in the fitnesses for hawks and doves gives p[t+1] = p[t]*(1+p[t](V-C)/2+(1-p[t]) V)/Wbar, where Wbar is the mean fitness. This can be simplified to p[t+1] = (1 + V - p[t](V+C)/2) p[t]/Wbar. In turn, Wbar can be simplified to Wbar = p[t]*Wh+(1-p[t])*Wd = 1 + V/2 - p[t]2 C/2
(b) First, solve for the change in p per generation, 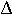 p = p[t+1] - p[t] = p[t] (1-p[t])(V/2 - p[t] C/2)/(1 + V/2 - p[t]2 C/2). Next, let the time frame shrink to an amount t, which shrinks both the gains and the costs to an amount proportional to t. The change in the frequency of hawks over the time interval t then becomes p[t] (1-p[t])(t V/2 - p[t] t C/2)/(1 + t V/2 - p[t]^2 t C/2). Finally, divide by t and take the limit as t goes to zero to get: dp/dt = p[t] (1-p[t])(V/2 - p[t] C/2).
p = p[t+1] - p[t] = p[t] (1-p[t])(V/2 - p[t] C/2)/(1 + V/2 - p[t]2 C/2). Next, let the time frame shrink to an amount t, which shrinks both the gains and the costs to an amount proportional to t. The change in the frequency of hawks over the time interval t then becomes p[t] (1-p[t])(t V/2 - p[t] t C/2)/(1 + t V/2 - p[t]^2 t C/2). Finally, divide by t and take the limit as t goes to zero to get: dp/dt = p[t] (1-p[t])(V/2 - p[t] C/2).
(c) =0, =1, and =V/C
(b) It is reasonable to assume that both V (the value of the resource) and C (the cost of fighting) are positive. Therefore, p=V/C will lie between 0 and 1 only if V is less than C.
(d) First expand the differential equation: f(p) = 1/2 p V - 1/2 p2 V - 1/2 p2 C + 1/2 p3 C.
Then take the derivative of this function with respect to the variable p: f'(p) = 1/2 V - p V - p C + 3/2 p2 C.
Then plug in the equilibrium =V/C: f'() = 1/2 V - (V/C) V - (V/C) C + 3/2 (V/C)2 C.
Then simplify: f'() = 1/2 V - V2/C - V + 3/2 V2/C = - 1/2 V + 1/2 V2/C = 1/2 V (V/C - 1)
The equilibrium will be stable when f'() < 0. We can assume that V (the value of the resource) is positive, so the equilibrium will be stable when V/C - 1 < 0, in other words when V < C. If V > C, then the equilibrium is unstable. Note that, from part (b), the equilibrium is stable whenever it is biologically valid (i.e. when V is less than C).
(e) When V>C, the internal equilibrium is unstable (and not biologically valid). Examining the payoff matrix when V>C, hawks are fitter than doves whether competing against a hawk (middle column) or against a dove (last column). Because hawks are always fitter, they spread to fixation regardless of the frequency of doves.
 Back to Biology 301 home page.
Back to Biology 301 home page.