
On July 1 1858, essays written by Charles Darwin and Alfred Russell Wallace describing evolution by natural selection were jointly read to the Linnean Society of London.
But let the external conditions of a country alter...Now, can it be doubted, from the struggle each individual has to obtain subsistence, that any minute variation in structure, habits, or instincts, adapting that individual better to the new conditions, would tell upon its vigour and health? In the struggle it would have a better chance of surviving; and those of its offspring which inherited the variation, be it ever so slight, would also have a better chance.-- Charles Darwin (1858)
-- Wallace (1858)
Evolution occurs whenever a population of individuals changes over time.
Evolution by natural selection occurs whenever
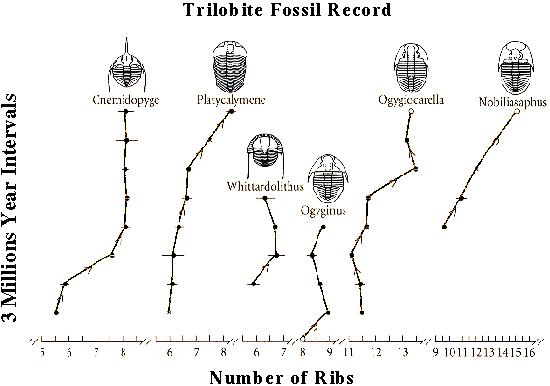
One can track evolutionary changes through the fossil record...
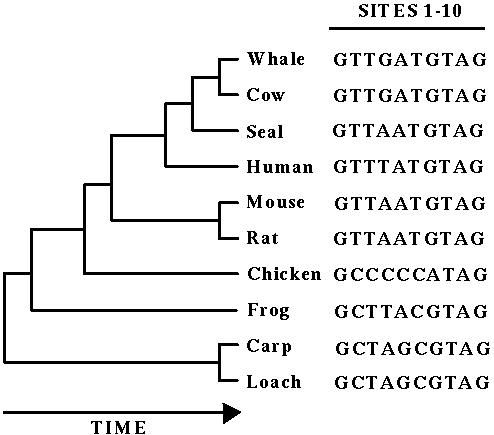
or through DNA...
Evolutionary rates vary widely over time, among species, and among traits.
Haldane (1949) proposed the darwin to measure evolutionary rates, with one darwin equaling a 2.718-fold change in a trait over one million years.
(GRAPH OF EVOLUTIONARY RATES)
Some species have changed rapidly in appearance over tens to hundreds of thousand years.
Cichlids in the African rift lakes
Columbine flowers
Sticklebacks in freshwater lakes of British Columbia
Sculpins in Lake Baikal (Siberia)
Drosophilid flies on Hawaii
Others have remained similar in appearance over tens to hundreds of million years.
Sequoia (redwoods)
Lungfish
Tuataras
Crocodiles
Monotremes (duckbills and echidnas)
Although these slowly changing species are called examples of "primitive" species and of "arrested evolution", many of them initially evolved rapidly and settled upon a successful way of life that is remarkably tolerant of change and disruption.
What factors limit rates of evolutionary change?
Faster rates of morphological evolution happen in rapidly changing environments and in relatively "empty" environments.
New islands (e.g. Galapagos, Hawaii), land and lakes following glacial retreat (e.g. BC, Lake Baikal), and new rifts lakes (e.g. Lake Victoria) pose both new environmental challenges and allow evolution "room to maneuver".
Darwin's finches of the Galapagos islands and the honeycreepers of the Hawaiian islands have undergone rapid morphological evolution in large part because the islands are relatively free of competitors.
In fairly static environments, the rate of evolution slows down.
(GRAPH OF RESPONSE TO SELECTION FROM LENSKI AND TRAVISANO, 1994)
Evolution can only go where mutations lead it.
There must be some traits that never appear through mutation, e.g. wheels with axles.
Yet, mutations do not appear to limit evolution strongly, in the sense that the immense diversity in rates of morphological evolution does not correlate with rates of evolution at the DNA level.
Indeed, turn-over rates in a gene (substitutions) are fairly constant across a variety of taxa.
(Comparisons of
Nevertheless, in especially harsh or novel situations, the rate of evolution must depend to some extent on the appearance of appropriate mutations (e.g. antibiotic resistance).
A recent experiment by de Visser et al. (1999) using asexual bacteria has shown that mutations can be limiting, especially in novel environments (i.e. low in glucose) and in small populations.
(GRAPH FROM DE VISSER)
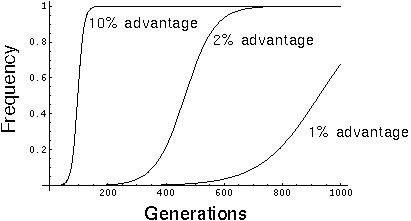
Although differential survival and reproduction cause changes to a population, these changes are not instantaneous.
It can take thousands of generations for a rare gene (say 1/10000) to become prevalent due to selection:
Furthermore, many new mutations may be lost before they even have a chance to spread.
Selection is less effective in small populations than in large populations. Consequently, one of the risks faced by small populations is that they will be unable to evolve fast enough to a changing environment. In such cases, the inefficiency of natural selection limits the rate of evolution.
Linkage - A gene can be advantageous and spread in a population when considered on its own, but genes are never on their own.
For example, imagine that red coloration is currently favourable and a new mutation occurs at one of many colour genes:
By chance, this mutation happened in an individual with many blue genes and few red genes. Even though the mutation is favorable, it will likely be lost unless sex occurs and mixes up the different genes.
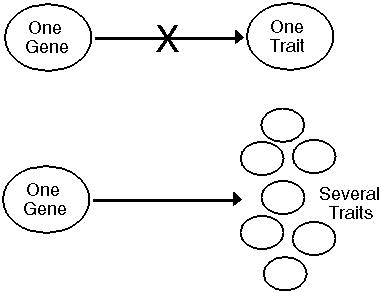
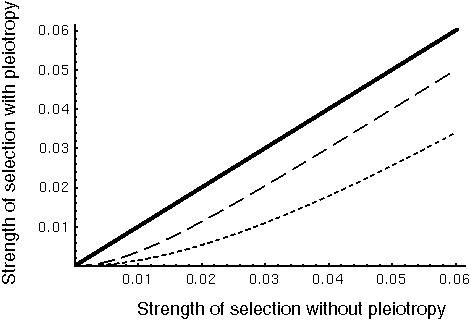
Pleiotropy - Genes often play multiple roles.
A mutation that does a wonderful new thing will not necessarily spread if it disrupts other things that a gene does.
These pleiotropic constraints on genes will reduce their response to selection and will slow the overall rate of evolutionary change.
Number of genes - When genes are highly constrained in their function, the rate of evolution may be increased by duplicating genes or whole genomes.
Gene duplication has played an important role in evolution.
EXAMPLE: Homeotic genes
Whole scale genome duplication may also free up genes to evolve novel functions.
EXAMPLE: In plants, groups with polyploidy have more species than related taxa (in 18/22 cases), which may be due to increased rates of evolution following genomic doubling.










Environment

Mutations

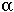 -globin genes from various vertebrates, Kimura 1983.)
-globin genes from various vertebrates, Kimura 1983.)
Efficiency of Selection
Architectural constraints
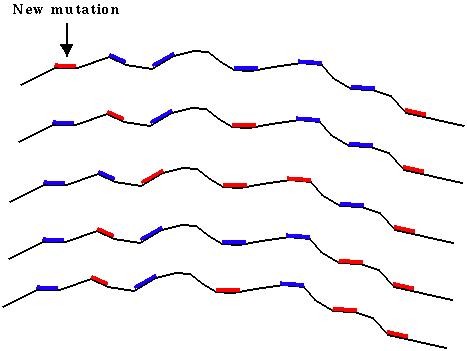
 Sex frees us from our baggage.
Sex frees us from our baggage.
Conclusion
...all limit evolution to different degrees in different times and places.