|
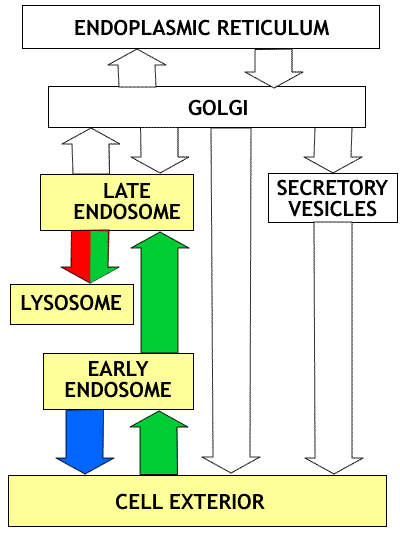
Overview of the endocytotic pathway: The compartments involved are highlighted in yellow. Proteins taken into the cell by receptor mediated endocytosis are transferred to early endosomes (green arrow) where receptors are removed and returned to the cell surface (blue arrow). The proteins are then moved to late endosomes (green arrow) and finally to lysosomes (green arrow) where digestion occurs. The lysosomes receive both proteins from the cell surface (green) and digestive enzymes coming from the Golgi (red). Click on figure to enlarge. |
We are dealing with two related phenomena:
In phagocytosis bacteria or other large particles are taken into membrane limited vesicles. Phagocytosis occurs in response to activation of surface receptors. This results in the opening of calcium channels, and the increase in the internal Ca++ concentration triggers the formation of a phagosome or food vacuole. Phagocytic cells play an important role in removal of foreign organisms (bacteria) and in the scavenging of dead or damaged cells.
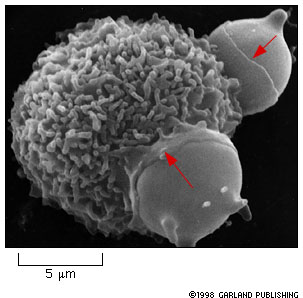 |
Figure 14-28. Macrophage engulfing two red blood cells. Notice
the collar-like pseudopodium creeping over the surface of the red blood
cells (red arrows). Given what you know about the cytoskeleton, what do
you think is happening inside the macrophage to make this happen?
The book says that macrophages eat 1011 red blood cells in each of us each day!!! That is a lot of cells! This occurs in the spleen. |
Pinocytosis. This involves the uptake of macromolecules and fluid by cells. The macromolecules bind to the cell surface. This is followed by the formation of a vesicle containing the surface membrane with the attached molecules and the surrounded fluid. Macrophages, for example ingest about 25% of their own volume in fluid each hour and an area of membrane equal to its surface every 30 minutes. There has to be a substantial traffic in vesicles fusing with the plasma membrane to maintain a steady state. This give you some idea of how important the constitutive secretory pathway is! Pinocytosis takes place via the formation of clathrin coated pits and vesicles. The clathrin and related proteins are shed from the endocytotic vesicles after they are formed and are recycled in the formation of new vesicles. The membrane vesicles eventually fuse with endosomes, and the macromolecular material they contain is digested.
The binding of macromolecules to proteins at the cell surface triggers a process known as receptor mediated endocytosis. The concentration of bound molecules triggers the formation of a vesicle. Macromolecules may be concentrated up to 1000-fold by accumulation on the cell surface before incorporation into vesicles. The concentration of cargo proteins occurs through the process discussed earlier in the lecture on vesicle formation; cargo molecules bind to receptors molecules that then bind to adaptin and coat protein (clathrin, in this case) that result in their concentration in the area of the forming membrane bud that will give rise to the endocytotic vesicle.
Endocytosed macromolecules are sorted in Early Endosomes. Early endosomes are relatively small vesicles that receive membrane and cargo from endocytotic vesicles. Early endosomes are main sorting sites on the endocytotic pathway, as the trans Golgi network serves this function in the secretory pathway. Endosomes receive a number of different types of receptors along with their cargo. Early endosomes have an acidic internal environment due to proton pumps in their membrane. The acidic environment causes many receptors or carrier proteins to release their cargo of bound macromolecules.
Most of the receptors are sorted out and returned to the plasma membrane by vesicle transport. In a well studied case with LDL receptors (see below) a receptor makes one round trip into the cell and back every 10 minutes.
 Fig. 14-29. Click to enlarge. |
Receptor-mediated endocytosis of Low Density Lipoprotein (LDL).
|
Macromolecules that are removed from their receptors are transferred to lysosomes for digestion. Most of the receptors are returned to the same plasma membrane domain (e.g. basal or apical) from which they came, others are passed to lysosomes and are destroyed. Still others, with their cargo are transferred to another plasma membrane domain and eventually transfer their cargo from one extracelular space to another, e.g. from the distal to the lateral face of an epithelial cell as shown in the diagram below.
|
Fig. 14-30. Click to enlarge. |
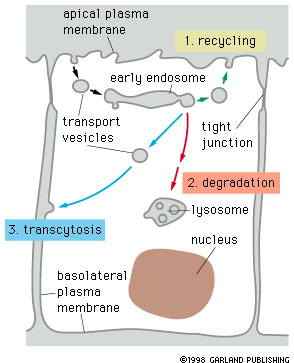
Three possible fates of receptor proteins involved in
receptor mediated endocytosis.
Receptors that are not specifically retrieved from early endosomes are transferred to lysosomes and degraded. Retreived receptors are returned either to the membrane domain of origin (recycling) or to another domain (transcytosis). If the ligand bound to the receptor stays bound in the acidic environment of the endosome, it will follow the same pathway as the receptor, otherwise it will be transferred to a lysosome and degraded. |
Macromolecules from early endosomes that are destined for degradation in lysosomes are passed in vesicles to Late Endosomes. Late endosomes also receive vesicles containing lysosomal enzymes from the trans Golgi network. The mannose-6-phosphate receptors that carry the lysosomal enzymes separate from their cargo in the acidic environment of the late endosome and are recycled to the trans Golgi network. The lysosomal enzymes and the macromolecules from early endosomes are then transferred by vesicles to lysosomes.
These processes are summarized in the following diagram:
 |
Recycling of cell surface receptors and mannose- 6 - phosphate receptors by endosomes. The green circles represent cargo molecules coming from formation of endocytotic vesicles at the cell surface. The red squares represent lysosomal enzymes carried from the trans Golgi network to the endosome by mannose-6 -phosphate receptors. Both types of receptors are recycled. Transport vesicles carry both lysosomal enzymes and macromolecules to be degraded to lysosomes. |
Sorting processes in cells are not perfect and some lysosomal proteins end up in vesicles that are secreted. Cells have a retrieval pathway to catch these errant lysosomal enzymes. The lysosomal enzymes bind to mannose- 6- phosphate receptors at the plasma membrane, the proteins are brought to endosomes and sent on to lysosomes while the receptors are returned to the trans-Golgi network. This rather elaborate system is simply a consequence of the normal operation of the sorting pathways. The mannose-6-phosphate receptors are at the surface because the targeting process that brings most of them back to the trans Golgi network is also not perfect.
There is a very nice description of the sorting and assembly of lysosomal components on pages 474 - 477 in the text. You should read this material carefully and be able to trance a molecule of a lysosomal enzyme from its point of origin to its final site of action.
Lysosomes are the digestive system of cells. These vesicles are a specialized set of secretory vesicles produced by the Golgi and contain a mixture of some 40 types of digestive enzymes, including those that degrade nucleic acids, proteins and lipids. All of these enzymes have optimum activity at about pH 5. This acid pH is maintained in lysosomes, as in endosomes, by proton pumps in the membrane. The membrane of the lysosome is resistant to action of its own digestive enzymes due to the extensive glycosylation of the proteins on the lumen side of the membrane.